Image Analysis Pipeline
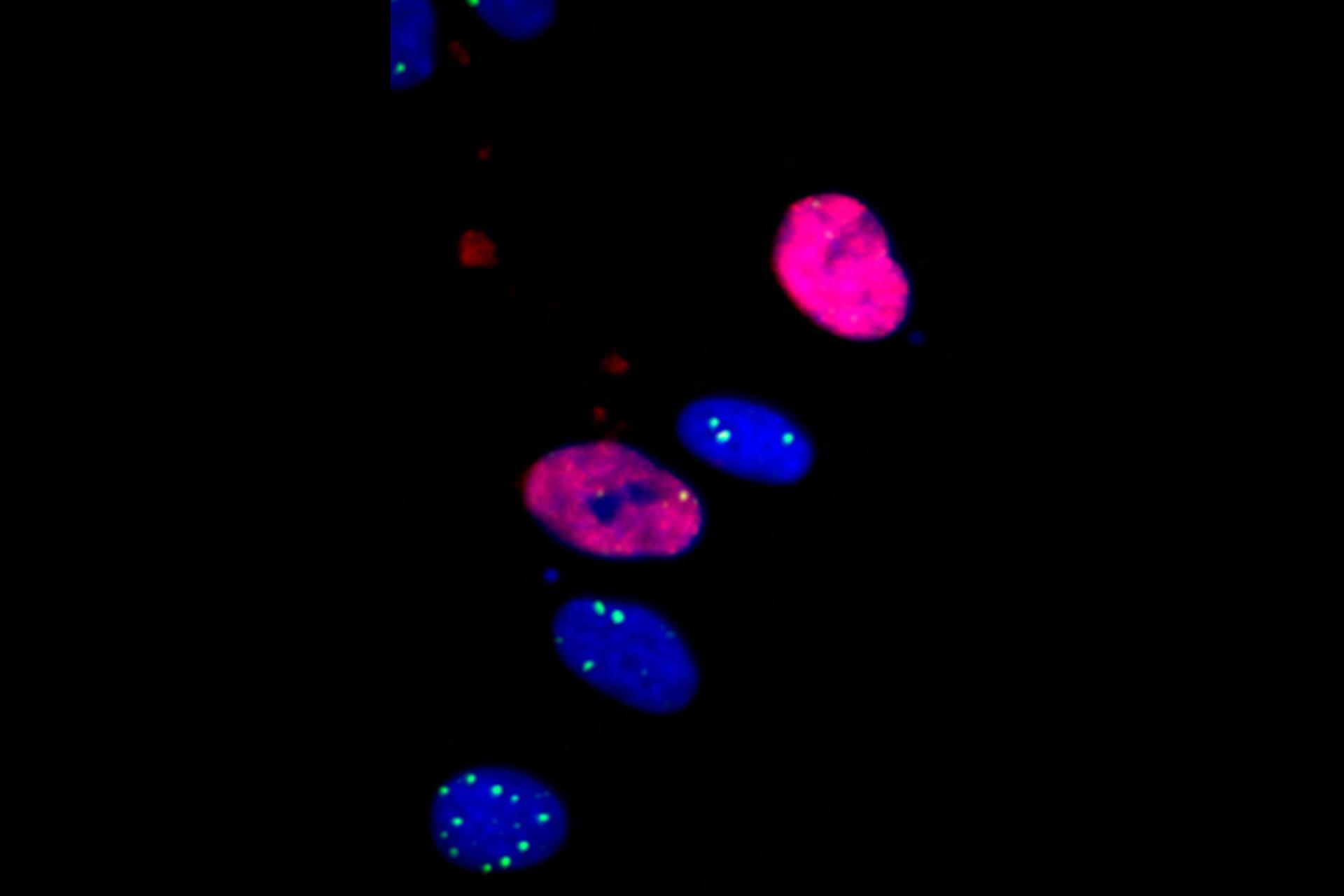
In a first step, both DAPI and 53BP1 channels were background subtracted to remove any background that might interfere with the detection of micronuclei, nuclei and DNA-damage foci. Especially micronuclei and DNA-damage foci especially can have small sizes and low intensities, making them hard to segment without good pre-processing
In the DAPI channel, we next detected both nuclei and micronuclei via watershed segmentation and separated both object groups by size and intensity (micronuclei are a factor of 100 smaller and are less intense than nuclei). The distance function was used to determine the distance from nuclei to the nearest micronucleus, which allowed later stratification of nuclei into micronucleus-positive and -negative fractions based on a distance cutoff.
By employing cut-off filters for DAPI and EdU channel intensity, nuclei were then further separated into three cell cycle phases: G0/G1 phase nuclei with DAPIlow/EdUlow total nuclear intensities, S-phase nuclei with EdUhigh and G2/M phase nuclei with DAPIhigh/EdUlow.
DNA-damage foci were derived also with watershed segmentation. Nuclei and raw foci were set into parent-child relationship by the compartment operation. This ensured that only DNA-damage foci within segmented nuclei were considered and allowed later stratification of DNA-damage foci along cell cycle phases.
This complete image processing pipeline is available under “Pipeline.xml” (part of the case study files download at the end of the webpage). We invite you to download a free trial of ZEISS arivis Pro and experience it in the software. Find all links at the bottom of this page.